Abstract
Human persistence as a species depends on biological reproduction, yet survival and flourishing also rely on numerous “non‑persistent” systems that do not themselves reproduce but are essential to those who do. This article examines how persistence is constructed at two levels: (a) biological systems, in which gamete production, fertilization, gestation, and birth are supported by neural circuits and evolutionary pressures that generate self‑preservation and sexual drives; and (b) social systems, in which families, communities, and institutions assemble both reproductive and non‑reproductive roles into integrated structures that allow populations to endure. After outlining basic reproductive biology and the neural bases of survival and sexual motivation, the article analyzes how modern technologies—such as assisted reproductive techniques—decouple sexual intimacy from conception while still requiring sperm and egg at the cellular level. It then turns to “non‑persistent” entities: somatic cells, sterile individuals, non‑reproductive vocations, and infrastructural roles that terminate in themselves but support the continuity of lineages and societies. Treating these as components of larger persistent systems clarifies how value and purpose are distributed beyond genes alone. The article concludes by raising normative questions: given that only a small fraction of individuals are strictly necessary for biological persistence, how should societies recognize and organize the contributions of those whose lives are non‑reproductive or non‑persistent, yet indispensable to the ongoing human project (e.g., caregiving, teaching, institutional work).
**Keywords:** human reproduction, persistence, non‑persistent systems, survival instinct, sexual behavior, assisted reproduction, social roles, evolutionary biology, family structures, generativity, ethics of reproduction
1. Introduction
Human life persists because new humans keep being born, raised, and integrated into social worlds that can sustain them. At a basic biological level, this requires the joining of sperm and egg, successful gestation, and viable offspring who themselves may eventually reproduce (Alberts et al., 2015). At a social level, however, persistence depends on a much wider architecture: people who never have children nonetheless work, teach, heal, protect, and care in ways without which the next generation could not survive or flourish (Hrdy, 2009).
This dual reality raises several questions. What exactly is the relation between individual bodies, sex, and species‑level persistence? How do biological systems embed both reproductive and non‑reproductive components into coherent wholes? And how do societies assemble “non‑persistent” lives and roles—those that do not extend themselves genetically—into structures that still contribute to long‑term survival and meaning? This paper proposes a framework for thinking about these questions, distinguishing between biological persistence and broader forms of generativity, and arguing that non‑persistent systems are integral rather than peripheral to the human story (Erikson, 1963; Sober & Wilson, 1998).
2. Biological Foundations of Persistence
2.1 Reproduction and the life cycle
In humans, biological persistence requires: (a) gametogenesis, the production of haploid gametes (sperm in testes, eggs in ovaries); (b) fertilization, the fusion of sperm and egg to form a diploid zygote; and (c) gestation and birth, involving embryonic and fetal development in a uterus culminating in live birth (Alberts et al., 2015). While not every individual must reproduce, a population must maintain sufficient fertility and child survival for the lineage to continue. From a population‑genetic perspective, the species persists when, over time, enough individuals successfully transmit genes to new generations to offset mortality and genetic drift (Frankham et al., 2014).
2.2 Neural bases of survival and sexual drives
At the organism level, survival and reproduction are supported by conserved neural circuits. Brainstem and hypothalamic systems regulate vital functions (respiration, circulation, thermoregulation) and coordinate rapid responses to threats (fight, flight, freeze) (LeDoux, 2012). Limbic structures such as the amygdala detect danger cues and trigger defensive states, while dopaminergic pathways and basal ganglia mediate motivation, reward, and habit formation (Berridge & Kringelbach, 2015; Panksepp, 1998).
Sexual behavior involves interactions among hypothalamic nuclei, limbic structures, and cortical regions, influenced by hormones and sensory input (Georgiadis & Kringelbach, 2012). These systems do not operate in isolation; they are embedded in networks for learning and prediction (including hippocampal and default‑mode activity), enabling organisms to acquire and refine strategies for finding mates, avoiding harm, and navigating social environments (Andrews‑Hanna et al., 2014).
2.3 Evolutionary pressures and the will to persist
Evolution by natural selection favors traits that enhance differential survival and reproduction (Mayr, 2001). Early self‑replicating chemistries that failed to maintain their structure, protect themselves, or produce copies were naturally eliminated, while those with improved self‑maintenance and reproductive reliability proliferated (Szostak, 2017). Over long timescales, this cumulative filtering yields organisms whose physiology and behavior are strongly oriented toward persistence—what is often experienced subjectively as a “will to live” or a drive to protect offspring and kin (Schopenhauer, 1966; Sober & Wilson, 1998).
This “will” does not require conscious intention. It is an emergent property of systems in which configurations that do not preserve themselves vanish from evolutionary history, leaving only those that do (Dawkins, 1976).
3. Assisted Reproduction and the Partial Decoupling of Sex and Conception
3.1 Technologies of conception
Modern reproductive technologies have significantly expanded the pathways by which sperm and egg can meet. In vitro fertilization (IVF) and intracytoplasmic sperm injection (ICSI) enable fertilization outside the body and can overcome various forms of infertility (Johnson, 2019). Intrauterine insemination (IUI) allows sperm to be placed in the reproductive tract without coitus. Donor sperm, donor eggs, and gestational surrogacy distribute genetic, gestational, and social parenthood across different individuals (Inhorn & Birenbaum‑Carmeli, 2008).
These techniques loosen the once‑tight coupling between sexual intercourse and conception, permitting reproduction in the absence of heterosexual coitus and sometimes in the absence of any sexual act at all between the eventual social parents.
3.2 What remains non‑negotiable
Despite this decoupling, certain biological constraints remain. Human reproduction still depends on the existence of viable sperm and viable eggs, and on embryonic development in a suitable gestational environment (currently a human uterus, though experimental work on ectogestation is ongoing) (Kaposy & Lotz, 2019). Early development remains sensitive to biological and environmental factors, including maternal health, intrauterine conditions, and early caregiving (Gluckman & Hanson, 2006).
Thus, technology can alter the social and interpersonal context of reproduction, but it does not eliminate the underlying requirement for gametes, gestation, and development.
3.3 Implications for persistence
These developments mean that persistence is no longer tied solely to the sexual behavior of heterosexual couples. Single parents, same‑sex couples, and others can have genetically related or intended children through assisted means (Inhorn & Patrizio, 2015). At the same time, reproductive capacity becomes more dependent on complex technical, legal, and economic infrastructures, which themselves must persist if the population is to maintain fertility options (Franklin & Roberts, 2006).
4. Non‑Persistent Biological Systems
4.1 Somatic cells and organism‑level persistence
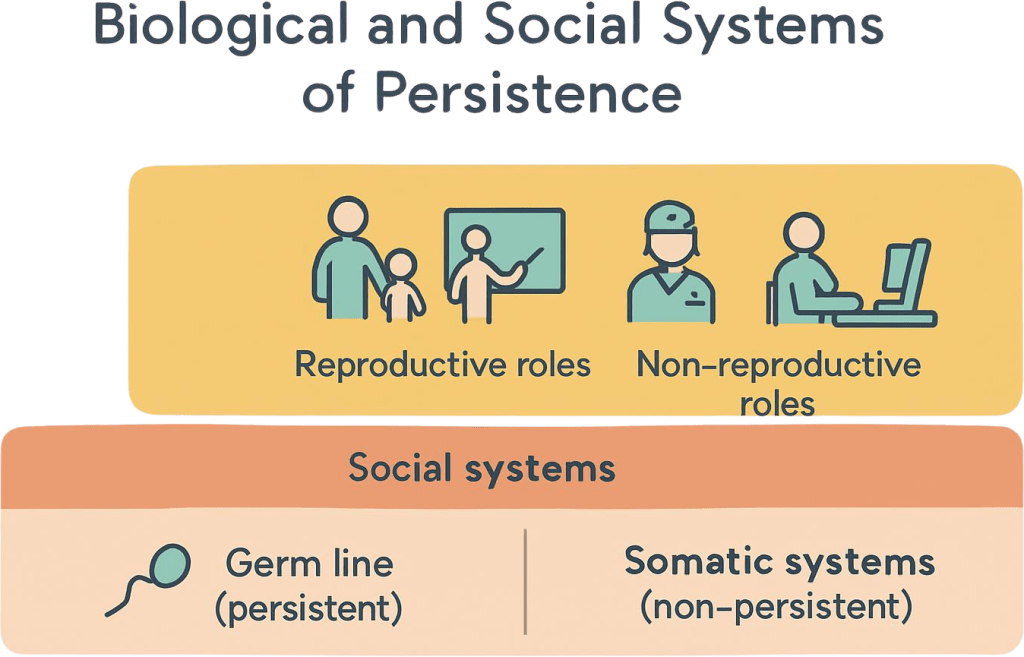
Within individual organisms, most cells are “non‑persistent” at the lineage level: somatic cells divide, perform their functions, and eventually die without transmitting genetic material to the next generation. Only germ cells—those that become gametes—contribute directly to genetic persistence (Gilbert & Barresi, 2016). Yet somatic cells are essential; they form tissues, organs, and systems that keep the organism alive long enough to reproduce and care for offspring. In multicellular life, a vast number of non‑reproductive cells exist precisely to support the reproduction and survival of the organism and its germ line.
4.2 Sterility, infertility, and non‑reproductive organisms
At the organism level, many individuals across species are sterile, infertile, or do not reproduce for ecological, social, or behavioral reasons. In some systems (such as eusocial insects), whole castes are structurally non‑reproductive but crucial for colony persistence (Wilson, 1971). In humans, infertility, celibacy, and child‑free lives are common and often chosen or embraced rather than simply suffered (Park, 2002).
Such individuals can still contribute to genetic and cultural persistence by aiding kin (inclusive fitness), supporting community members, or strengthening institutions that protect offspring survival (Hamilton, 1964; Hrdy, 2009). Evolutionary models show that non‑reproductive helpers can be favored when their contributions sufficiently increase the reproductive success of related individuals or the stability of the group (Sober & Wilson, 1998).
5. Social Assemblies of Persistent and Non‑Persistent Roles
5.1 Family systems
In human families, reproductive adults and children are embedded in networks that include non‑reproductive individuals: extended kin, household members, and community caregivers. Anthropological and developmental research shows that “alloparents” such as grandparents, aunts, uncles, and older siblings often play crucial roles in child survival and socialization (Hrdy, 2009). These networks distribute the labor of raising children, managing households, and maintaining emotional and material stability. From a systems perspective, the family is a composite of persistent and non‑persistent roles that together support generational continuity.
5.2 Community and institutional structures
Beyond the family, persistence depends on broader structures: education systems that transmit knowledge and skills; economic systems that organize production, distribution, and work; health systems that reduce mortality and increase lifespan; and legal and political systems that regulate conflict and allocate resources (Giddens, 1984). People working in these systems—many of whom may not reproduce—are indispensable to the survival prospects of the population. Their roles are non‑persistent at the genetic level but crucial at the societal level.
5.3 Non‑persistent vocations and generativity
Many vocations can be understood as non‑persistent in the narrow sense (they do not directly generate offspring) but generative in a wider sense: they create and sustain conditions under which others can live and reproduce. Teaching, caregiving, scientific research, infrastructure maintenance, artistic creation, and governance all fall into this category (McAdams & de St. Aubin, 1992). These roles illustrate a key point: persistence is not only about copying genes but also about maintaining the material, informational, and relational structures that make survival and flourishing possible.
6. Bodies, Sex, and Social Meaning
6.1 Sex beyond reproduction
Human sexual behavior serves multiple functions besides procreation. It fosters bonding and intimacy, offers pleasure, communicates affection or commitment, and sometimes acquires spiritual or ritual significance (Bancroft, 2009). While reproductive capacity depends on gametes and gestation, sexual scripts vary widely and are shaped by culture, orientation, identity, and technology (Gagnon & Simon, 2005). Many sexual relationships are consciously non‑reproductive and yet deeply meaningful.
6.2 Family diversity and persistent outcomes
Diverse family forms—including single‑parent households, blended families, queer families, communal and chosen families—can all support child development and contribute to persistence, provided they have access to resources and supportive structures. Empirical literature suggests that stability, safety, and responsive care matter more for child outcomes than specific family configurations (Biblarz & Stacey, 2010). Recognizing this diversity highlights that the link between bodies, sex, and persistence is mediated by social recognition, legal protections, and economic conditions (Cherlin, 2010).
7. Ethical and Conceptual Implications
7.1 Value beyond genetic contribution
If only a small fraction of humans are strictly required to sustain the population biologically, a narrow valuation that privileges reproduction alone would undervalue vast numbers of lives and roles. An ethical perspective attentive to non‑persistent systems must affirm the worth of individuals regardless of reproductive status, recognize non‑reproductive contributions as central to human persistence and flourishing, and question social arrangements that marginalize or exploit non‑reproductive labor while benefiting from it (Kittay, 1999; Nussbaum, 2011).
7.2 Persistence and the question of purpose
Understanding how persistence works biologically and socially does not by itself answer what persistence is for. Different worldviews answer that question differently—invoking divine purposes, liberation from suffering, humanist ideals, or existential self‑creation (Cottingham, 2003; Taylor, 1989). Whatever answer one adopts, it must grapple with the fact that persistence is a cooperative achievement of both persistent and non‑persistent systems.
8. Conclusion
Bodies, sex, and persistence are tightly intertwined, but not in a simple or singular way. At the biological level, human persistence requires the joining of sperm and egg, gestation, and successful development of offspring. At the neural and evolutionary level, survival and sexual drives arise from circuits and histories that favor self‑maintenance and reproduction. At the same time, societies are built from many non‑persistent components—cells that do not transmit genes, individuals who do not reproduce, and roles that terminate in themselves yet support the conditions under which others can live and bear children.
Recognizing this layered structure allows a more accurate and humane account of what keeps humanity going. It invites ethical reflection on how to honor contributions beyond reproduction and how to design social systems that distribute both the burdens and benefits of persistence more justly.
9. Future Work
Future work could explore several directions. First, quantitative models could estimate how non‑reproductive roles (by profession, caregiving, etc.) influence population survival, health, and resilience. Second, cross‑cultural studies could examine how different societies value and organize non‑persistent roles, and how these configurations affect demographic stability and well‑being. Third, neurobiological research could investigate how brain systems for reward, attachment, and meaning are engaged by non‑reproductive forms of care and contribution. Fourth, ethical and legal analyses could assess policies affecting reproductive and non‑reproductive individuals (e.g., parental leave, support for caregivers, assisted reproduction access) in light of their role in persistence. Finally, speculative work on technological futures—artificial gestation, gamete derivation from somatic cells, radical lifespan extension—could assess how emerging capabilities might reconfigure the balance between persistent and non‑persistent systems, and what new responsibilities they create.
References
Alberts, B., Johnson, A., Lewis, J., Raff, M., Roberts, K., & Walter, P. (2015). *Molecular biology of the cell* (6th ed.). Garland Science.
Andrews‑Hanna, J. R., Smallwood, J., & Spreng, R. N. (2014). The default network and self‑generated thought: Component processes, dynamic control, and clinical relevance. *Annals of the New York Academy of Sciences, 1316*(1), 29–52.
Bancroft, J. (2009). Human sexuality and its problems (3rd ed.). Churchill Livingstone.
Berridge, K. C., & Kringelbach, M. L. (2015). Pleasure systems in the brain. *Neuron, 86*(3), 646–664.
Biblarz, T. J., & Stacey, J. (2010). How does the gender of parents matter? *Journal of Marriage and Family, 72*(1), 3–22.
Cherlin, A. J. (2010). *The marriage‑go‑round: The state of marriage and the family in America today*. Vintage.
Cottingham, J. (2003). *On the meaning of life*. Routledge.
Dawkins, R. (1976). *The selfish gene*. Oxford University Press.
Erikson, E. H. (1963). *Childhood and society* (2nd ed.). W. W. Norton.
Frankham, R., Ballou, J. D., & Briscoe, D. A. (2014). *Introduction to conservation genetics* (2nd ed.). Cambridge University Press.
Franklin, S., & Roberts, C. (2006). *Born and made: An ethnography of preimplantation genetic diagnosis*. Princeton University Press.
Gagnon, J. H., & Simon, W. (2005). *Sexual conduct: The social sources of human sexuality* (2nd ed.). Aldine.
Georgiadis, J. R., & Kringelbach, M. L. (2012). The human sexual response cycle: Brain imaging evidence linking sex to other pleasures. *Progress in Neurobiology, 98*(1), 49–81.
Giddens, A. (1984). *The constitution of society: Outline of the theory of structuration*. University of California Press.
Gilbert, S. F., & Barresi, M. J. F. (2016). *Developmental biology* (11th ed.). Sinauer.
Gluckman, P. D., & Hanson, M. A. (2006). *Mismatch: Why our world no longer fits our bodies*. Oxford University Press.
Hamilton, W. D. (1964). The genetical evolution of social behaviour. I & II. *Journal of Theoretical Biology, 7*(1), 1–52.
Hrdy, S. B. (2009). *Mothers and others: The evolutionary origins of mutual understanding*. Harvard University Press.
Inhorn, M. C., & Birenbaum‑Carmeli, D. (2008). Assisted reproductive technologies and culture change. *Annual Review of Anthropology, 37*, 177–196.
Inhorn, M. C., & Patrizio, P. (2015). Infertility around the globe: New thinking on gender, reproductive technologies and global movements in the 21st century. *Human Reproduction Update, 21*(4), 411–426.
Johnson, M. (2019). Human in vitro fertilization: A brief history. *Methods in Molecular Biology, 2006*, 3–15.
Kaposy, C., & Lotz, M. (2019). The ethics of ectogenesis. *Bioethics, 33*(1), 28–34.
Kittay, E. F. (1999). *Love’s labor: Essays on women, equality, and dependency*. Routledge.
LeDoux, J. (2012). *Rethinking the emotional brain*. *Neuron, 73*(4), 653–676.
Mayr, E. (2001). *What evolution is*. Basic Books.
McAdams, D. P., & de St. Aubin, E. (1992). A theory of generativity and its assessment through self‑report, behavioral acts, and narrative themes in autobiography. *Journal of Personality and Social Psychology, 62*(6), 1003–1015.
Nussbaum, M. C. (2011). *Creating capabilities: The human development approach*. Belknap Press.
Panksepp, J. (1998). *Affective neuroscience: The foundations of human and animal emotions*. Oxford University Press.
Park, K. (2002). Stigma management among the voluntarily childless. *Sociological Perspectives, 45*(1), 21–45.
Schopenhauer, A. (1966). *The world as will and representation* (E. F. J. Payne, Trans.). Dover. (Original work published 1818)
Sober, E., & Wilson, D. S. (1998). *Unto others: The evolution and psychology of unselfish behavior*. Harvard University Press.
Szostak, J. W. (2017). The narrow road to the deep past: In search of the chemistry of the origin of life. *Angewandte Chemie International Edition, 56*(37), 11037–11043.
Taylor, C. (1989). *Sources of the self: The making of the modern identity*. Harvard University Press.
Wilson, E. O. (1971). *The insect societies*. Harvard University Press.

Leave a comment